
- Classification
- MYXINI
- MYXINIFORMES
- MYXINIDAE
-
Fish Classification
-
Class
MYXINI Hagfishes -
Order
MYXINIFORMES Hagfishes -
Family
MYXINIDAE Hagfishes -
Genera
Eptatretus(5)
Family MYXINIDAE
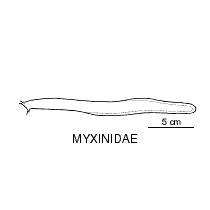
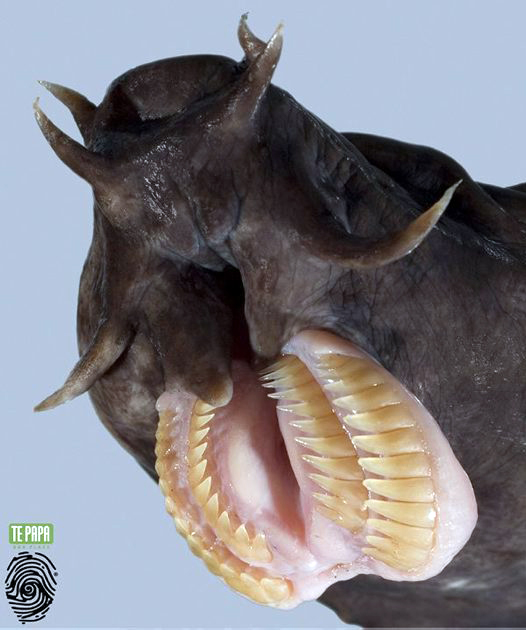
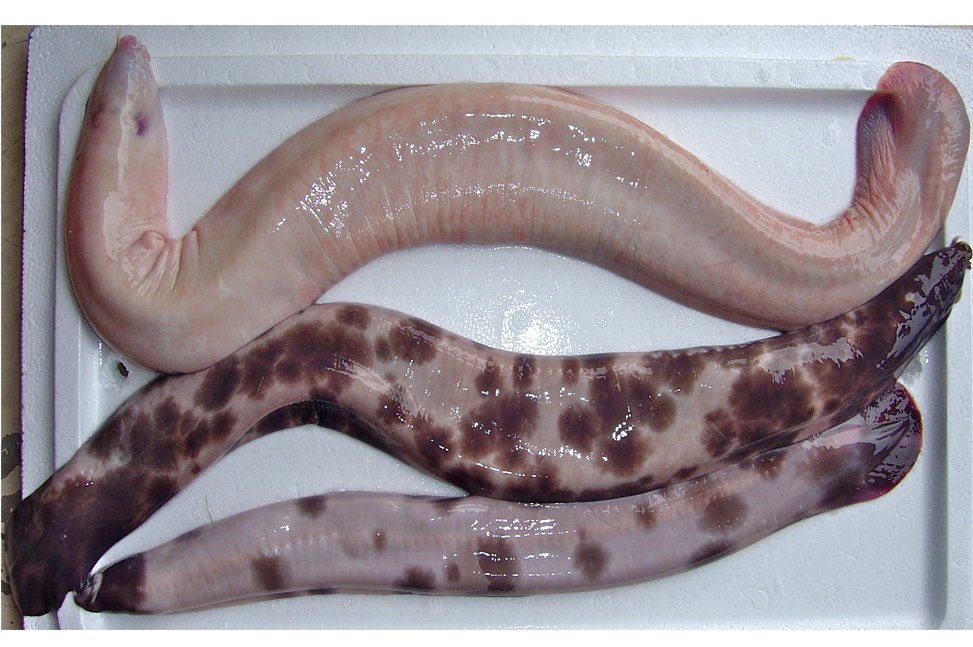
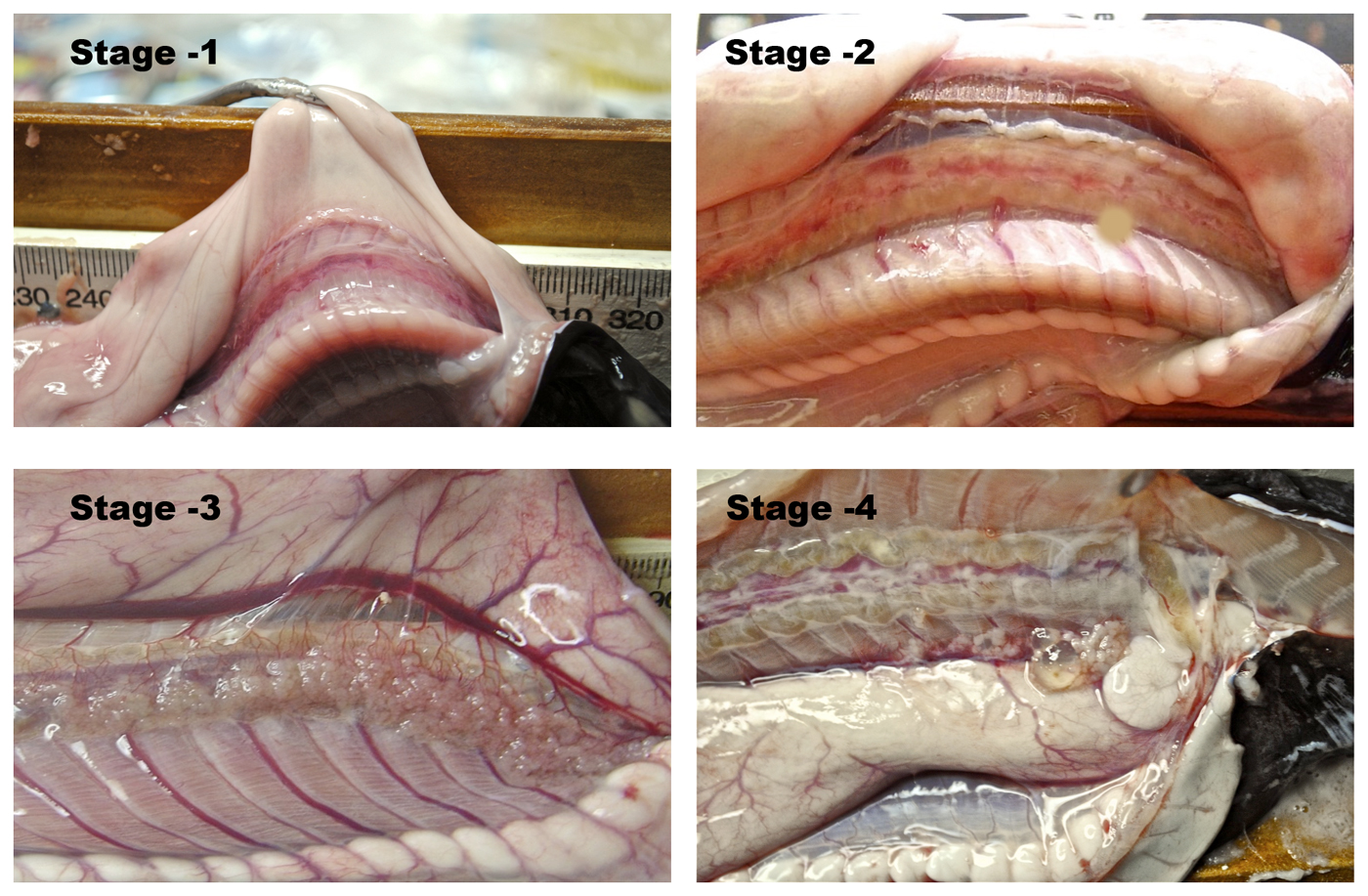
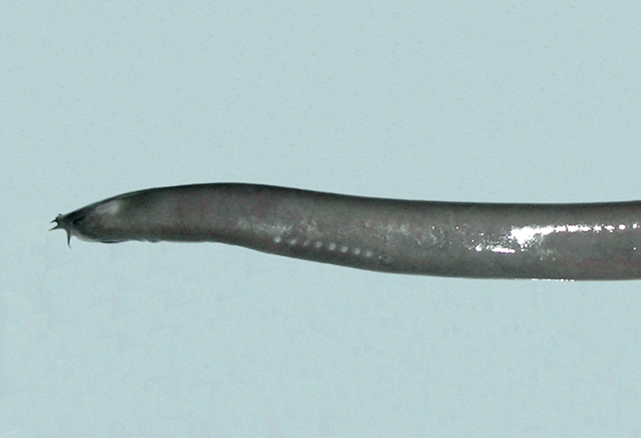
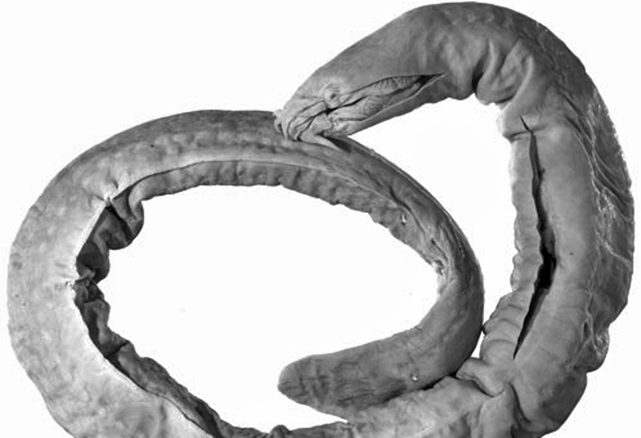
Primitive eel-like fishes found worldwide, mostly in temperate regions. They have naked eel-like bodies with 1-16 pairs of external gill openings on each side, no paired fins and no dorsal fin, and the caudal fin extending onto the upper and lower surface. Hagfish have degenerate eyes, a jawless mouth surrounding by barbels and horny rasping teeth on the tongue. They produce enormous amounts of slime to avoid predation.








More Info
|
Family Taxonomy |
The family comprises 7 genera and about 76 species separated into two subfamilies: Myxininae with one common gill opening on each side and Eptatretinae with 5-16 openings on each side. A single genus, Eptatretus, and four descibed species are known to occur Australian waters (Mincarone & Fernholm 2010). Relatively little material has been collected in Australia and additional species may occur here. Fossil hagfishes date back 300 million years ago. |
|
Family Distribution |
Hagfishes are marine bottom-dwelling fishes found in all oceans. Most species have an antitropical distribution and prefer to live on soft bottoms in cool temperate waters from six to 1300 metres. |
|
Family Size |
To 1.1 metres. |
|
Family Colour |
Generally greyish or brownish in colour. |
|
Family Feeding |
Hagfishes scavenge on dead or dying marine life, primarily on fishes and invertebrates, but also on marine mammals that have sunk to the ocean floor. They also prey on living fishes and invertebrates. They produce enormous amounts of an incredibly sticky slime to protect themselves by clogging up the mouths and gills of their predators. They feed by rasping flesh, often going completely inside sunken carcasses and feeding from the inside out. Recent studies have shown that hagfishes also 'feed' by absorbing nutrients through their permeable skin and gills. |
|
Family Reproduction |
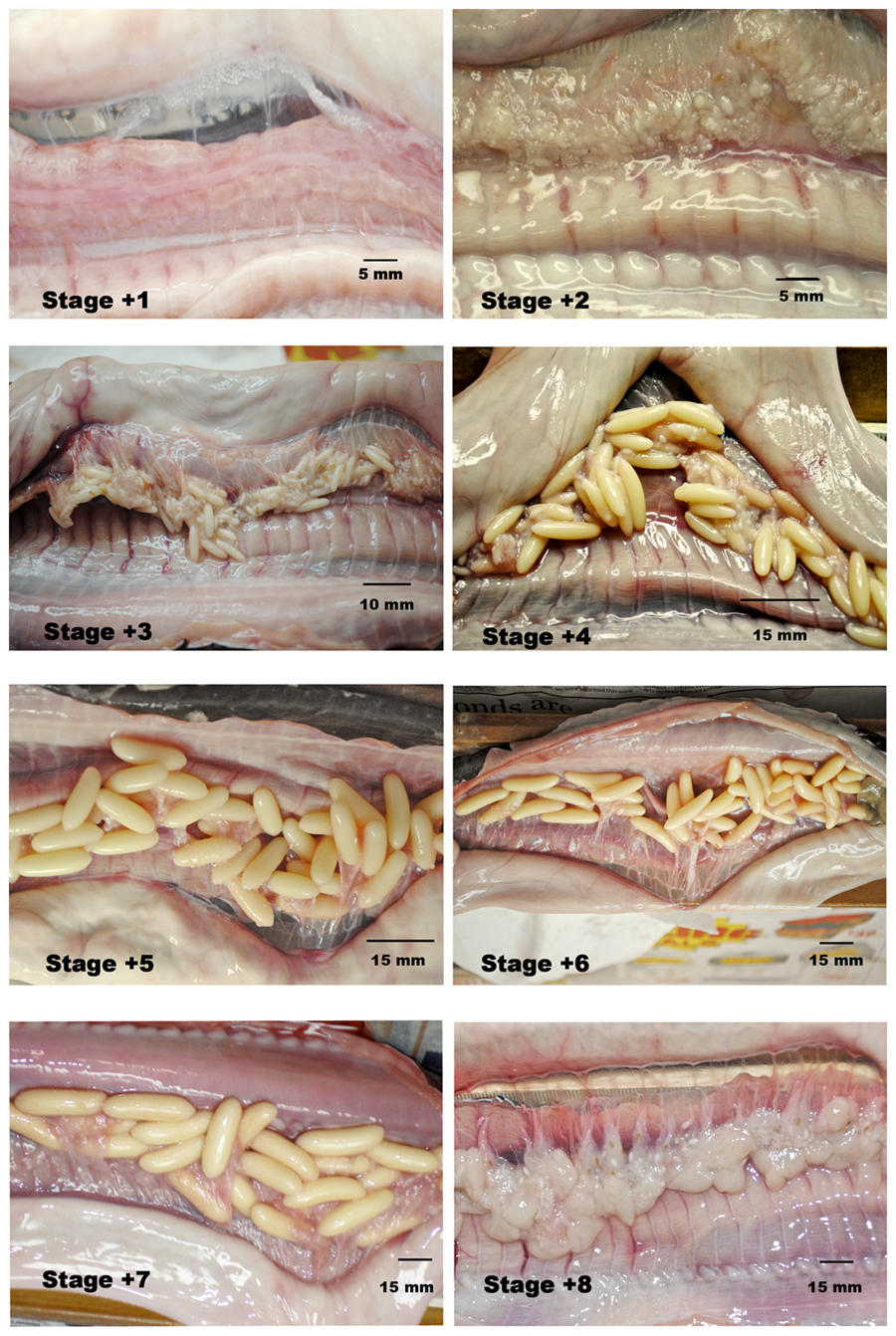
Both sterile and hermaphroditic individuals have been found. Spawning and fertilisation may occur in burrows. Hagfishes lay relatively few large eggs which they presumably attached to the substrate by filaments. There is no larval stage and young hagfishes develop directly from the egg. |
|
Family Commercial |
Although of no interest to Australian fisheries, hagfishes are commercially fished worldwide - slime eel fishing - and are heavily exploited in some regions. Not only are hagfishes consumed along with their eggs and slime, their skins are tanned, made into leather and marketed as "eelskin". Hagfish populations in some areas are threatened by overfishing. The copious amounts of slime they produce is problematic for commercial fishers using baited traps in deeper waters. Hagfishes are consumed as food in Aisa. |
|
Family Conservation |
Hagfishes have been recently assessed using the International Union for Conservation of Nature (IUCN) criteria, and 9 of the 76 known hagfish species were found to be threatened, including species found in southern Australia (Knapp et al. 2011). Threats include overfishing and habitat destruction. |
|
Family Remarks |
When threatened, hagfishes secrete a gelatinous slime containing mucus and thousands of intermediate filament protein threads that are manufactured within specialized gland thread cells. The protein threads are extremely strong and incredibly long, and are able to rapidly uncoil without becoming tangled. (Winegard et al. 2014) |
|
Author |
Dianne J. Bray |
References
Collin, S.P., Davies, W.L., Hart, N.S. and Hunt, D.M. (2009) The evolution of early vertebrate photoreceptors. Philosophical Transactions of The Royal Society B-Biological Sciences 364 1531: 2925-2940.
Donoghue, P.C.J., Forey, P.L. & Aldridge, R.J. 2000. Conodont affinity and chordate phylogeny. Biological Reviews of the Cambridge Philosophical Society 75: 191–251.
Fernholm, B. 1998. Hagfish systematics. P. 578, In Jorgensen, J.M., J.P. Lomholt, R.E. Weber & H. Make. (eds) The Biology of Hagfish. Chapman & Hall, London.
Fernholm, B. & K. Holmberg. 1975. The eyes in three genera of hagfish (Eptatretus, Paramyxine and Myxine) – a case of degenerative evolution. Vision Res. 15: 253–259.
Fudge, D., Levy, N., Chiu, S. & Gosline, J. 2005. Composition, morphology and mechanics of hagfish slime. J. Exp. Biol. 208: 4613–4625.
Glover, C.N., C. Bucking & C.M. Wood. 2011. Adaptations to in situ feeding: novel nutrient acquisition pathways in an ancient vertebrate. Proc. R. Soc. B doi:10.1098/rspb.2010.2784 (2011)
Gomon, M.F. & M. Mincarone. 2008. Myxinidae, In: Gomon, M.F., D.J. Bray & R.H. Kuiter (eds.) Fishes of Australia's southern coast. Reed New Holland & Museum Victoria.
Huang, K.F., Mok, H.K. & Huang, P.C. 1994. Hagfishes of Taiwan (II): Taxonomy as inferred from mitochondrial DNA diversity. Zool. Stud. 33: 186–191.
Jansson, H., Wyoni, P.I., Fernholm, B., Bredwad, M., Mierzykowaka, A. & Tegelstrom, H. 1995. Genetic relationships among species of hagfish revealed by protein electrophoresis. J. Fish. Biol. 47: 599–608.
Janvier, P. 2007. Evolutionary biology: born-again hagfishes. Nature 446: 622–623.
Jorgensen, J.M., J.P. Lomholt, R.E. Weber & H. Malte. (eds.) The Biology of Hagfishes. 1998. Chapman and Hall, New York. 578 p.
Knapp, L., M.M. Mincarone, H. Harwell, B. Polidor, J. Sanciangco & K. Carpenter. 2011. Conservation status of the world’s hagfish species and the loss of phylogenetic diversity and ecosystem function. Aquatic Conserv: Mar. Freshw. Ecosyst. 21: 401–411. http://onlinelibrary.wiley.com/doi/10.1002/aqc.1202/pdf
Martini, F.H. 1998. The ecology of hagfishes. p. 57-77. In: J.M. Jørgensen, J.P. Lomholt, R.E. Weber & H. Malte. The biology of hagfishes. Chapman & Hall, London. 578 pp.
Minarcone, M.M. 2009. Eptatretus cirrhatus. In: IUCN 2011. IUCN Red List of Threatened Species. Version 2011.1. . Downloaded on 18 August 2011. http://www.iucnredlist.org/apps/redlist/details/154825/0
Mincarone, M.M. & Fernholm, B. 2010. Review of the Australian hagfishes with description of two new species of Eptatretus (Myxinidae). Journal of Fish Biology 77: 779–801.
Mincarone, M.M. & Stewart, A.L. 2006. A new species of giant seven-gilled hagfish (Myxinidae: Eptatretus) from New Zealand. Copeia 2006: 225–229.
Near, T.J. 2009. Conflict and resolution between phylogenies inferred from molecular and phenotypic data sets for hagfish, lampreys, and gnathostomes. Journal of Experimental Zoology Part B: Molecular and Developmental Evolution 312B: 749–761.
Negishi, A., C.L. Armstrong, L. Kreplak, M.C. Rheinstadter, L-T. Lim , T.E. Gillis & D.S. Fudge. 2012. The production of fibers and films from solubilized hagfish slime thread proteins. Biomacromolecules 13: 3475–3482. PDF
Nelson, J.S. 2006. Fishes of the World. John Wiley & Sons, New Jersey, 4th Edn, 601 pp.
Ota KG, Kuraku S, Kuratani S. 2007 Hagfish embryology with reference to the evolution of the neural crest. Nature 446: 672-675.
Patzner, R.A. 1998. Gonads and reproduction in hagfishes, pp. 378-395. In: J.M. Jørgensen, J.P. Lomholt, R.E. Weber, & H. Malte (eds), The biology of hagfishes, Chapman & Hall, London.
Paxton, J.R., Gates, J.E. & Hoese, D.F. 2006. Myxinidae, pp. 38–39. In: Beesley, P.L. & Wells, A. (eds) Zoological Catalogue of Australia, Volume 35. Part 1. Collingwood: ABRS & CSIRO Publishing.
Powell ML, Kavanaugh SI, Sower SA (2005) Current knowledge of hagfish reproduction: implications for fisheries management. Integr Comp Biol 45(1): 158-165. doi:10.1093/icb/45.1.158.
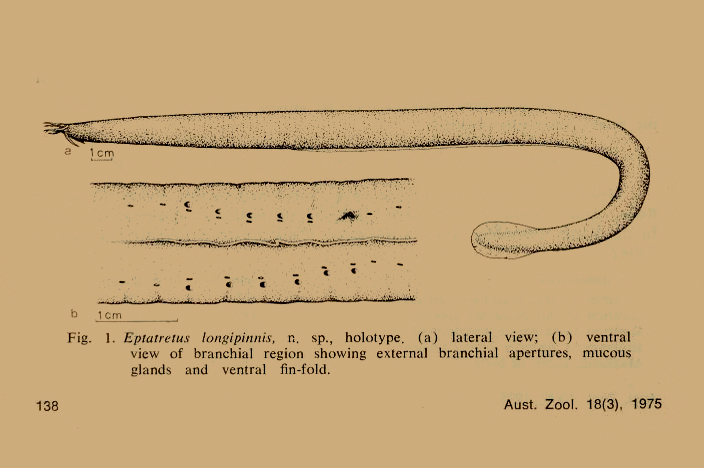
Strahan, R. 1975. Eptatretus longipinnis, n.sp., a new hagfish (family Eptatretidae) from South Australia, with a key to the 5 – 7 gilled Eptatretidae. Aust. Zool. 18(3): 137–148, figs. 1–2.
Winegard, T. & Fudge, D. 2010. Deployment of hagfish thread skeins requires the transmission of mixing forces via mucin strands. J. Exp. Biol. 213: 1235–1240.
Winegard, T., J. Herr, C. Mena, B. Lee, I. Dinov, D. Bird, M. Bernards, S. Hobel, B. Van Valkenburgh, A. Toga & D. Fudge. 2014. Coiling and maturation of a high-performance fibre in hagfish slime gland thread cells. Nature Communications 2014; 5 DOI: 10.1038/ncomms4534 Abstract
Yu, S., Zhang, W., Li, L., Huang, H., Ma, F. & Li, Q. 2008. Phylogenetic analysis of 48 gene families revealing relationships between Hagfishes, Lampreys, and Gnathostomata. Journal of Genetics and Genomics 35: 285–290.